




2. 内蒙古自治区林业科学研究院 呼和浩特 010000;
3. 中国科学院新疆生态与地理研究所 国家荒漠-绿洲生态建设工程技术研究中心 乌鲁木齐 830011;
4. 新疆策勒荒漠草地生态系统国家野外科学观测研究站 策勒 848300
2. Inner Mongolia Academy of Forestry Sciences, Hohhot 010000, China;
3. National Engineering Technology Research Center for Desert-Oasis Ecological Construction, Xinjiang Institute of Ecology and Geography, Chinese Academy of Sciences, Urumqi 830011, China;
4. Cele National Station of Observation and Research for Desert-Grassland Ecosystems, Cele 848300, China
位于新疆南疆的塔里木盆地是我国沙漠化最严重的地区之一。该区气候干旱,降水稀少,蒸发强烈,植被稀疏。这里的植物区系组成贫乏——各类草甸、灌丛和荒漠河岸林生长在低洼地和河流两岸。它们是一类典型的荒漠河岸植被,其生存主要依赖于地下水,构成了隐域性植被[1]。这些多年生植被不仅是保护绿洲的生态屏障,而且是当地重要的放牧场所,对当地畜牧业发展有着重要的贡献[2]。20世纪下半叶,由于人口的快速增加和过度开垦,荒漠-绿洲过渡带的多年生植物遭到了大面积的破坏,继而导致了风沙灾害,使得大面积耕地被流沙吞没,并产生了饲料短缺等严重问题[3]。因此,研究区域优势植物的生理生态学特性,揭示植物与水分的关系及稳定修复机制[4, 5],对退化荒漠生态系统的可持续经营具有重要意义。
位于新疆北疆的准噶尔盆地被公认为是对全球变化最敏感的地区之一[6]。该地区降雨量和蒸发量反差巨大,年均降水量仅为60—250 mm,年蒸发量则在2 000 mm以上;由于冬季有稳定的积雪覆盖,最大积雪厚度多在20 cm以上,早春融雪为荒漠植物的生长发育提供了有利的条件[7]。降水格局的变化会导致土壤水分时空分布的动态变化[8],影响到土壤种子库的萌发,进而补充和更新幼苗[9]。这些变化也必将影响植物的生长和生理生态特性[10],进而有可能影响到旱地生态系统的结构和功能属性[11]。因此,水分影响着荒漠生态系统的各个环节,以及荒漠区的植被恢复与重建[12, 13]。梭梭(Haloxylon ammodendron C. A. Mey.)是我国西北荒漠半荒漠地区的珍贵植物资源。梭梭集中分布区在准噶尔盆地,约占我国梭梭总面积的68.2%[14]。梭梭植被是农耕绿洲的天然屏障,具有不可估量的生态、经济和社会效益[15]。在气候变化和人类活动的影响下,准噶尔盆地天然梭梭林分布面积减小,盖度下降。保护干旱区生态环境,防止沙漠扩展,拯救、保护梭梭已迫在眉睫。
分布于内蒙古科尔沁沙地和浑善达克沙地的疏林草原植被,不仅是防风固沙、保护沙区生态环境和周边土地资源的一种重要的植被,也是耐旱沙生植物的重要物种基因库和草原野生动物的重要避难所和栖息地。天然疏林草原中榆树(Ulmus pumila L.)具有强抗旱性特征及较大遗传多样性变异潜力[16]。然而,近年来榆树群落正处于严重的退化状态,主要表现为天然更新严重不足[17]。这与过度放牧下牲畜啃食幼苗幼树、人类经济活动的过度干扰等造成榆树种子失去了萌发所需要的位置稳定性有关[18],当然也与气候变化导致生境恶化存在一定的联系。
2 我国不同典型区主要优势植被的稳定修复途径 2.1 新疆塔里木盆地塔里木盆地南缘区域降水极少,荒漠优势植物通过根系直接或间接与地下水相连通或从地下水毛细上升区获得持续稳定的水分来源,基于水势差调节、渗透调节、形态调整等多种调节方式来应对干旱胁迫,从而实现植物的稳定维持和有效修复。因此,维持区域地下水稳定是保障优势植物可持续经营的重要前提。
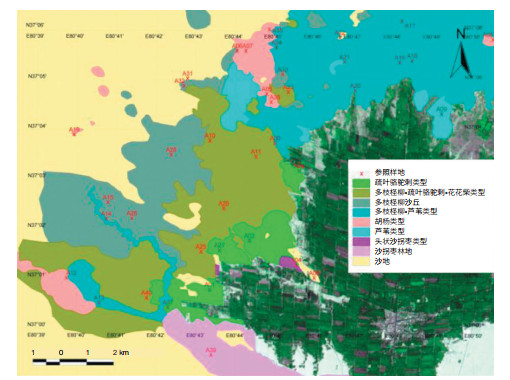
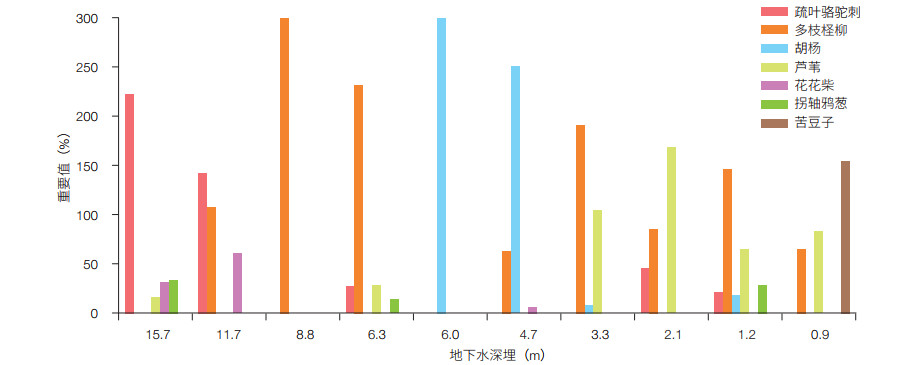
在塔里木盆地南缘荒漠-绿洲过渡带分布有5种优势植被类型(图 1):疏叶骆驼刺(Alhagi sparsifolia Shap.)、胡杨(Populus euphratica Oliv.)、多枝柽柳(Tamarix ramosissima Ledeb)、头状沙拐枣(Calligonum caput-medusae Schrenk)和芦苇(Phragmites australis Trin.)[2]。这些植物的分布特征和群落构成与地下水埋深密切相关。研究表明[19],在地下潜水埋深15.7 m地段,疏叶骆驼刺在群落中占有绝对优势,并且呈均匀分布状态;在地下水埋深8.8 m地段,植被由多枝柽柳形成的单一植物群落组成,但多枝柽柳的频度、相对多度和盖度值都非常低,群落中有大片空白地;在地下潜水埋深6.3 m地段,同样是以多枝柽柳为主的群落,但群落盖度有较大程度的增加、生物多样性升高。从群落的数量指标分析,多枝柽柳在群落中占有绝对优势的地位,但分布上并不均匀。在地下水埋深6.0—4.7 m地段,是胡杨优势群落。在地下水埋深4.5 m地段,群落的盖度和组成成分增加,多枝柽柳的密度甚至超过胡杨,但胡杨仍占绝对优势(图 2)。在地下水埋深3.3 m地段,多枝柽柳虽然在群落中占据明显优势,但密度下降,而芦苇的重要性明显上升。当地下水埋深升高到2.1 m,芦苇成为群落的主要成份。尽管群落中仍有多枝柽柳和疏叶骆驼刺出现,但重要性明显下降。
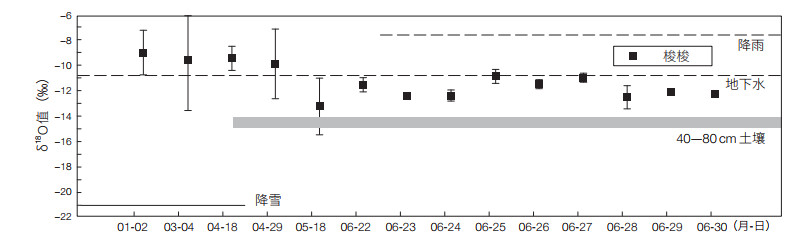
2.2 新疆准噶尔盆地新疆准噶尔盆地优势植物梭梭的维持水源具有多途径特点,地下水、融雪、降雨形成的浅层土壤水是其主要水源[20, 21]。处于不同林龄和不同生长时段的梭梭对不同水源利用形式的转换是维持其正常生存和生长的重要途径。研究表明,幼龄梭梭在每年4月主要利用浅层土壤水,利用比例为62%—95%;幼龄梭梭在每年5月主要利用0—50 cm土层的土壤水,利用比例达67.5%;成株梭梭在每年7月主要利用250—400 cm土层的土壤水,利用比例达94.2%[22]。在每年5—9月梭梭主要利用地下水,利用比例为68%—100%(图 3)。由冬季融雪和春季降水补给的浅层土壤水和地下水是梭梭种群可利用的2类重要水源,而梭梭对不同时期发生的2场相似量级的降水具有不同程度的响应。梭梭的水分利用动态反映了其对干旱环境的适应[23]。
2.3 内蒙古科尔沁沙地和浑善达克沙地实行季节性倒场放牧和草原承包制背景下的保护性利用,可有效恢复草地植被,实现草地的可持续经营利用[24, 25]。研究表明,近年来内蒙古西辽河流域科尔沁沙地和浑善达克沙地东部的榆树群落正处于严重的退化状态,主要表现为天然更新严重不足(图 4a)。封育保护对榆树自然落种更新十分有利,其存活曲线趋向于Deevey Ⅰ型(图 4b)。林下草地植被既是疏林草原的重要组成部分,也直接影响着榆树疏林的自然恢复。

|
| 图 4 内蒙古西辽河流域科尔沁沙地和浑善达克沙地的东部的榆树种群年龄结构(a)和种群存活曲线(b) |
此外,沙地半灌木蒿类植被是沙地植被中最有代表性的主体组成部分,它与生物气候条件的变化相适应并表现出明显的区域性特征,从半干旱到干旱区依次分布有科尔沁沙地的差巴嘎蒿(Artemisia halodendron Turcz.et Bess.)、浑善达克沙地的褐沙蒿(Artemisia intramongolica H.C.Fu)和毛乌素沙地向西深入到腾格里沙漠西缘的油蒿(Artemisia ordosica Krasch)等植被,在区域生态环境保护和农牧业生产方面发挥着极其重要的作用,维持其稳定具有十分重要的意义[26]。半灌木蒿类植被具有基本相同的生态习性和繁殖特点,种子表面的沙蒿胶,遇到水形成粘液可防止种子发生位移;半灌木蒿类幼苗和成年个体在沙埋和干旱胁迫下形成多种抗旱适应机制[27-29];这些植被也都是有性繁殖和无性繁殖的兼性克隆植物。这些适应性使得蒿类植被具有在干旱、多风沙环境下快速自我修复的能力。只要通过封育为退化半灌木蒿类植被提供稳定的环境条件,就可以有效地促进其恢复;适度放牧利用也是保持其群落稳定的关键;而对于退化严重的群落,还可采取平茬方式进行复壮更新[30]。
3 不同典型区主要优势植被的可持续管理技术 3.1 合理处理沙漠前沿植被与洪水和地下水的关系(1)合理处理洪水和沙漠前沿植被的关系。塔里木盆地南缘是降水极端稀少的区域,而且程度已经达到1 000年以来的最低点。植物繁殖需要良好的土壤水分条件,这不仅是种子萌发的需要,而且是保证幼苗根系达到地下水的需要。由于水分缓慢地渗过土壤淤积层,短时间(几天内)的洪水不会引起深层土壤水分的充分饱和。所研究的植物都没有出现远离现代河床、形成大面积分布范围的现象[3],甚至在土壤表面到毛细管水边缘距离超过20 m的立地也没有出现[31]。
(2)稳定维持地下水和前沿植被的关系。在塔里木盆地沙漠周缘的沙丘中,从沙丘顶端到地下水之间的距离主要是通过植物的根系来连接的,还可以通过(植物)的主干和粗枝来连接的。植物的生产力、木质部水势和δ13C值随着植物离地下水距离的增加而降低,这表明(植物体内)水分传输的阻力和水分在木质部中的传输距离有关[31, 32]。沙漠前沿的多年生植物不仅能够而且需要和地下水相联系,因此前沿植被的存在依赖于持久的地下水供给[33]。
(3)对沙漠前沿植被的收获量不应超过植被的增长量[34]。植被的死亡和更新要达到平衡。在确定植被年生长量的基础上,可以计算植被的允许利用强度。为了避免浪费灌溉水,在植被更新措施中,应运用能促进植物根系尽快生长而达到地下水的灌溉方法。当植物根系达到地下水以后,灌溉即可停止。为了在植被更新中确保灌溉水的节约利用,即使在土壤表面离地下水距离很远的立地,也需要调查选择合适的灌溉方法和研究植物根系对土壤水分的依赖程度,为实现植被的“近自然”恢复重建奠定技术基础。
3.2 充分利用积雪融水培育幼苗和免灌造林(1)利用雪墒播种幼苗培育关键技术。准噶尔盆地边缘冬季多年平均积雪厚度为20—30 mm,可以利用好的雪墒飞播或者人工辅助恢复梭梭。人工种植技术关键为:一次供水量为20 mm,沙埋厚度为1.0 cm。
(2)梭梭免灌造林技术。在准噶尔盆地,种植幼苗要利用好早春的积雪融水,造林应在每年3月底前完成。由于干旱区降水量低,通过改造微地形,形成积雪沟和积水坑的方法,让降水再分配,形成梭梭集水造林免灌植被造林方法。梭梭林合理种植密度为2.5 m×3 m,即1 333株/hm2作为初植密度和成林密度是较为合适的。其能够达到水分供需平衡,在不灌溉情况下,仅靠降水不会出现固沙林衰亡的结果[22]。
3.3 封育保护是促进优势植被稳定修复的先决条件封育保护对于防止人畜对沙漠(沙地)自然植被的破坏,以及提供稳定的环境条件具有十分重要的作用,也是促进自然植被稳定修复的先决条件。沙漠(沙地)自然植被的生理生态、生长繁殖与演替规律是其稳定修复与可持续经营的理论基础。系统性地开展自然植被方面相关的基础理论研究,对于制定自然植被保护、修复、经营和利用的技术措施和相关政策等,都具有十分重要的意义,因此应给予足够的重视和长期的支持。
3.4 统筹兼顾生态用水、扶持区域经济发展,也将为沙漠前沿主要自然植被的有效保育和合理利用创造重要条件在21世纪水资源需求总量不断增加的大背景下,应把生态用水摆在落实可持续发展的高度,与生产生活用水一视同仁,通盘考虑,合理分配上、中、下游用水,绿洲建设与荒漠治理用水。同时,通过节水满足农业规模扩大的需要,为保护和改造荒漠植被提供合理的生态灌溉水源。
| [1] |
张立运, 夏阳. 塔克拉玛干沙漠南缘生态环境的特点及治理问题. 干旱区资源与环境, 1994, 8(1): 72-79. |
| [2] |
张希明, Michael Runge. 塔克拉玛干沙漠边缘植被可持续管理的生态学基础. 北京: 科学出版社, 2006.
|
| [3] |
Bruelheide H, Vonlanthen B, Jandt U, et al. Life on the edge-To which degree does phreatic water sustain vegetation in the periphery of the Taklamakan Desert. Applied Vegetation Science, 2010, 13(1): 56-71. DOI:10.1111/j.1654-109X.2009.01050.x |
| [4] |
Zeng F J, Bleby T M, Landman P A, et al. Water and nutrient dynamics in surface roots and soils are not modified by shortterm flooding of phreatophytic plants in a hyperarid desert?. Plant and Soil, 2006, 279(1-2): 129-139. DOI:10.1007/s11104-005-0498-2 |
| [5] |
Li X Y, Zhang X M, Zeng F J, et al. Water relations on Alhagi sparsifolia in the southern fringe of Taklamakan Desert. Acta Botanica Sinica, 2002, 44(10): 1219-1224. |
| [6] |
Intergovernmental Panel on Climate Change. Summary for policymakers//Field C B, Barros V R, Dokken D J, et al (Eds). Climate Change 2014-Impacts, Adaptation and Vulnerability. Part A: Global and Sectoral Aspects. Cambridge: Cambridge University Press, 2014: 1-32.
|
| [7] |
王雪芹, 张元明, 蒋进, 等. 古尔班通古特沙漠南部沙垄水分动态——兼论积雪融化和冻土变化对沙丘水分分异作用. 冰川冻土, 2006, 28(2): 262-268. DOI:10.3969/j.issn.1000-0240.2006.02.017 |
| [8] |
Sponseller R A. Precipitation pulses and soil CO2 flux in a Sonoran Desert ecosystem. Global Change Biology, 2007, 13(2): 426-436. DOI:10.1111/j.1365-2486.2006.01307.x |
| [9] |
Vile D, Pervent M, Belluau M, et al. Arabidopsis growth under prolonged high temperature and water deficit:Independent or interactive effects?. Plant, Cell & Environment, 2012, 35(4): 702-718. |
| [10] |
Guérin M, Martin-Benito D, von Arx G, et al. Interannual variations in needle and sapwood traits of Pinus edulis branches under an experimental drought. Ecology and Evolution, 2018, 8(3): 1655-1672. DOI:10.1002/ece3.3743 |
| [11] |
Berdugo M, Delgado-Baquerizo Manuel, Soliveres S, et al. Global ecosystem thresholds driven by aridity. Science, 2020, 367: 787-790. DOI:10.1126/science.aay5958 |
| [12] |
刘国军, 张希明, 吕朝燕. 梭梭自然更新与维持生态学. 北京: 科学出版社, 2019.
|
| [13] |
赵文智, 程国栋. 干旱区生态水文过程研究若干问题评述. 科学通报, 2001, 46(22): 1851-1857. DOI:10.3321/j.issn:0023-074X.2001.22.002 |
| [14] |
陈昌笃, 张立运, 胡文康. 古尔班通古特沙漠的沙地植物群落、区系及其分布的基本特征植物. 生态学与地植物学丛刊, 1983, 7(2): 89-99. |
| [15] |
Kang J J, Duan J J, Wang S M, et al. Na compound fertilizer promotes growth and enhances drought resistance of the succulent xerophyte Haloxylon ammodendron. Soil Science and Plant Nutrition, 2013, 59(2): 289-299. DOI:10.1080/00380768.2012.763183 |
| [16] |
刘果厚, 贾宝丽. 浑善达克沙地榆遗传多样性的研究. 干旱区资源与环境, 2003, 17(5): 123-128. DOI:10.3969/j.issn.1003-7578.2003.05.024 |
| [17] |
李钢铁, 姚云峰, 邹受益, 等. 科尔沁沙地榆树疏林草原及其封育更新研究. 干旱区资源与环境, 2004, 18(4): 152-157. DOI:10.3969/j.issn.1003-7578.2004.04.031 |
| [18] |
李永庚, 蒋高明, 高雷明, 等. 人为干扰对浑善达克沙地榆树疏林的影响. 植物生态学报, 2003, 27(6): 829-834. DOI:10.3321/j.issn:1005-264X.2003.06.016 |
| [19] |
Li X Y, Lin L S, Zhao Q, et al. Influence of groundwater depth on species composition and community structure in the transition zone of Cele oasis. Journal of Arid Land, 2010, 2(4): 235-242. |
| [20] |
Liu G J, Lv C Y, Zhang X M, et al. Effect of water supply and sowing depth on seedling emergence in two Haloxylon species in the Jungar basin. Pakistan Journal of Botany, 2015, 47(3): 859-865. |
| [21] |
吕金岭, 张希明, 吕朝燕, 等. 准噶尔盆地南缘荒漠区梭梭维持水源初步研究. 中国沙漠, 2013, 33(1): 110-117. |
| [22] |
李银芳. 梭梭人工林密度研究. 中国沙漠, 1998, 18(1): 22-26. DOI:10.3321/j.issn:1000-694X.1998.01.004 |
| [23] |
戴岳, 郑新军, 唐立松, 等. 古尔班通古特沙漠南缘梭梭水分利用动态. 植物生态学报, 2014, 38(11): 1214-1225. |
| [24] |
姚雪玲, 李龙, 王锋, 等. 放牧方式对浑善达克沙地榆树疏林退化的影响. 生态学报, 2020, 40(5): 1663-1671. |
| [25] |
张文军, 王晓江, 邢旗, 等. 内蒙古干旱半干旱区植被恢复与可持续经营研究. 呼和浩特:内蒙古人民出版社, 2014, 168-206. |
| [26] |
郭柯. 毛乌素沙地油蒿群落的循环演替. 植物生态学报, 2000, 24(2): 243-247. DOI:10.3321/j.issn:1005-264X.2000.02.020 |
| [27] |
赵哈林, 曲浩, 赵学勇, 等. 差巴嘎蒿幼苗对沙埋的生态适应和生理响应. 生态学报, 2014, 34(20): 5832-5839. |
| [28] |
Li C P, Xiao C W. Above- and belowground biomass of Artemisia ordosica communities in three contrasting habitats of the Mu Us desert, Northern China. Journal of Arid Environments, 2007, 70(2): 195-207. DOI:10.1016/j.jaridenv.2006.12.017 |
| [29] |
李鑫豪, 闫慧娟, 卫腾宙, 等. 油蒿资源利用效率在生长季的相对变化及对环境因子的响应. 植物生态学报, 2019, 43(10): 889-898. DOI:10.17521/cjpe.2019.0214 |
| [30] |
王庆锁, 陈仲薪, 史振英. 油蒿草场的保护与改良. 生态学杂志, 1995, 14(4): 54-57. DOI:10.3321/j.issn:1000-4890.1995.04.009 |
| [31] |
Gries D, Zeng F J, Foetzki A, et al. Growth and water relations of Tamarix ramosissima and Populus euphratica on Taklamakan desert dunes in relation to depth to a permanent water table. Plant, Cell & Environment, 2003, 26(5): 725-736. |
| [32] |
Arndt S K, Arampatsis C, Foetzki A, et al. Contrasting patterns of leaf solute accumulation and salt adaptation in four phreatophytic desert plants in a hyperarid desert with saline groundwater. Journal of Arid Environments, 2004, 59(2): 259-270. DOI:10.1016/j.jaridenv.2004.01.017 |
| [33] |
Vonlanthen B, Zhang X M, Bruelheide H. On the run for water-Root growth of two phreatophytes in the Taklamakan Desert. Journal of Arid Environments, 2010, 74(12): 1604-1615. DOI:10.1016/j.jaridenv.2010.07.004 |
| [34] |
Gries D, Foetzki A, Arndt S K, et al. Production of perennial vegetation in an oasis-desert transition zone in NW Chinaallometric estimation, and assessment of flooding and use effects. Plant Ecology, 2005, 181(1): 23-43. |